
Строение и уровни организации днк. Структура молекулы днк Строение и свойства молекулы днк

Жизнь ДНК (дезоксирибонуклеиновых кислот)
Определение понятия "ДНК"
Ген - это совокупность сегментов ДНК, обуславливающих образование либо молекулы РНК, либо белкового продукта (Сингер М., Берг П., 1998).
У человека около 30000 генов. Во всём объёме ДНК структурные гены (т.е. те, которые кодируют белки, идущие на построение стуктур организма) занимают лишь 3-10%.
Наименьшая функциональная единица ДНК состоит из следующих элементов: структурный ген, регуляторные зоны, регуляторные гены.
Строение молекулы ДНК
Молекулы ДНК имеют вид длинных двойных цепей полимеров – полинуклеотидов, состоящих из мономеров – нуклеотидов. Двойная цепь закручена в спираль. Поэтому ДНК похожа на винтовую лестницу (посмотрите на рисунок вверху). Каждый нуклеотид включает одно из четырех азотистых оснований – аденин (А), гуанин (Г), цитозин (Ц) или тимин (Т), одну молекулу пентозы (пятиуглеродный сахар) и один остаток фосфорной кислоты. Обычно молекула ДНК состоит из двух комплементарных нитей, которые образуют двойную спираль. При этом аденин одной нити находится в паре с тимином другой (стабилизируется двумя водородными связями), а гуанин аналогично связан с цитозином (тремя водородными связями). Последовательность азотистых оснований в молекуле ДНК несет информацию, необходимую для синтеза белков. ДНК - очень длинные молекулы, состоящие из множества нуклеотидов. Например, геном человека состоит из 46 хромосом, основу которых составляют молекулы ДНК, которые в совокупности собраны примерно из 3 млрд нуклеотидны пар.
У эукариот генетический материал находится в ядре клетки в хромосомах. Хромосомы в активном состоянии существуют в виде хроматина. Хроматин содержит около 40% ДНК, 40% гистонов (щелочных белков), около 20% негистоновых хромосомных белков и немного РНК.
Видео: Строение хромосомы
ДНК мы можем отнести к "живым системам", к "живым молекулам" на том основании, что они лежат в основе жизни вообще, а также обладают рядом важнейших свойств живого, в частности, способностью к размножению. ДНК насктолько самостоятельны и самодостаточны, что способны существовать даже вне клетки - в виде вирусов. В своей жизни молекулы ДНК проходят жизненные этапы, напоминающие нам жизнь более сложных биологических систем - живых организмов. Это такие этапы как рождение, созревание, работа (деятельность) и "смерть".
Тема: Строение ДНК
Домашнее задание
- Знать и уметь писать структурные формулы нуклеотидов: А, Т, Г, Ц, У.
- Знать устройство молекул ДНК и их организацию в хромосомы.
- Знать способы связывания нуклеотидов в ДНК по вертикали и горизонтали. Понятие о 3"-5" связях.
- Уметь пользоваться таблицей генетического кода для построения молекул пептидов на основе участка ДНК размером от 12 и более нуклеотидов.
Видео: Хромосомы, митоз, репликация
Этапы жизни молекулы ДНК
Рождение (репликация) - созревание (хромосомы) - работа (транскрипция) - управление (регуляция) - видоизменение (мутация) - "смерть"
1. Репликация ДНК - рождение новой дочерней нити ДНК на родительской нити.
2. Созревание ДНК - формирование хромосомы.
3. Транскрипция ДНК - работа ДНК в виде матричного синтеза на ней РНК.
4. Регуляция транскрипции - управление деятельностью ДНК по транскрипции.
5. Репарация ДНК - восстановление повреждённых участков.
6. Изменения структуры ДНК - мутации, транспозоны.
7. Деградация ДНК - разрушение при каждом цикле репликации.
1. Рождение - репликация
Репликация ДНК проходит очень просто, на счёт "раз, два, три", то есть в три этапа: 1) инициация, 2) элонгация, 3) терминация.
1. Инициация - начинание
Мишень для запуска репликации
Репликация огромной молекулы ДНК начинается с возникновения репликативной точки. Эта точка имеет специфическую последовательность богатую парами А-Т. Такие учкастки в ДНК как раз и являются мишенями для белков, инициирующих репликацию. Именно к ним присоединяются специальные распознающие белки, которые обеспечивают присоединение ферментов репликации хеликазы
и топоизомеразы
(гиразы) и таким образом запускают процесс репликации. Хеликаза
расплетает ДНК на две цепи. Образуется репликативная вилка. Молекула ДНК жестко закреплена на ядерном матриксе и не может свободно вращаться при расплетании какого-либо участка. Это блокирует продвижение хеликазы по цепи. Топоизомераза надрезает нити ДНК и снимает структурное напряжение.
В одной репликативной вилке действуют две хеликазы, которые движутся в противоположных направлениях. Разделенные цепи фиксируются ДНК- связывающими белками. Участки формирования репликативной вилки называются «точками ori» (origin - начало). У эукариот одновременно образуется тысячи таких вилок, что обеспечивает высокую скорость репликации.
2. Элонгация - продолжение (удлиннение)
Наращивание дочерних цепей ДНК на двух родительских цепях происходит неодинаково. ДНК- полимераза III прокариот и δ- или α-ДНК-полимеразы эукариот могут осуществлять синтез новой цепи ДНК лишь в направлении 5’>3’, т.к. могут присоединить новый нуклеотид только к углероду в положении 3’, но не в положении 5’.
Цепь с такой направленностью называется лидирующей . На ней синтез дочерней нити ДНК идёт непрерывно. ДНК-полимераза III или δ-полимераза непрерывно присоединяют к ней комплементарные нуклеотиды.
Цепь с полярностью 3’>5’ является отстающей и достраивается по частям (также в направлении 5’>3’). α-ДНК-полимераза (или ДНК-полимераза III) синтезирует на этой цепи короткие участки - фрагменты Оказаки.
Синтез фрагментов Оказаки и лидирующей цепи начинается с образования РНК-праймеров (затравок ) длиной 10-15 рибонуклеотидов ферментом праймазой (РНК-полимеразой). Ни одна из ДНК-полимераз не способна начать синтез ДНК с нуля, а может лишь достраивать существующую цепь. Параллельно с образованием лидирующей цепи или фрагментов Оказаки происходит удаление рибонуклеотидов из праймеров и замена их нуклеотидами ДНК. Замена рибонуклеиновых участков (праймеров) на участки ДНК происходит с помощью β-ДНК-полимеразы, которая имеет как экзонуклеазную, так и полимеразную активность.
Таким образом, репликация невозможна без частичной временной транскрипции.
Скорость репликации (элонгации) ДНК составляет около 45000 нуклеотидов в минуту, таким образом, родительская вилка расплетается со скоростью 4500 об/мин. Это сопоставимо, например, со скоростью вращения охлаждающего вентилятора в компьютере (1300-4800 об/мин).
3. Терминация - завершение (окончание)
Завершение репликации происходит тогда, когда пробелы между фрагментами Оказаки заполнятся нуклеотидами (при участии ДНК-лигазы) с образованием двух непрерывных двойных цепей ДНК и когда встретятся две репликативные вилки. Затем происходит закручивание синтезированных ДНК с образованием суперспиралей.
Правильность репликации обеспечивается точным соответствием комплементарных пар оснований и действием ДНК-полимераз, которые обладают кроме полимеразной, еще и экзонуклеазной активностью и способны распознавать и исправлять ошибки. Если включается некомплементарный нуклеотид, то фермент делает шаг назад, отщепляет его и продолжает полимеразную реакцию. Поэтому процесс репликации является высокоточным.
После завершения репликации происходит метилирование ДНК в участках –GАТС- по аденину (с образованием N-метиладенина) и остаткам цитозина с образованием 5-метилцитозина. Метилирование не нарушает комплементарности цепей и является необходимым для формирования структуры хромосом и регуляции транскрипции генов.
У прокариот, таких как бактерии, ДНК способна реплицироваться, не распрямляясь в линейную молекулу, то есть оставаясь в характерной для неё кольцевой форме.
Видео: Репликация (удвоение) ДНК
2. Созревание - формирование хромосомы и хроматина
3. Работа - транскрипция
Видео: Блокировка работы гена
4. Управление - регуляция
5. Восстановление (починка) - репарация
6. Видоизменение - мутация .
7. "Смерть" - деградация при репликации.
DNA Logic - это технология ДНК-вычислений, которая сегодня находится в зачаточном состоянии, однако в будущем на нее возлагаются большие надежды. Биологические нанокомпьютеры, вживляемые в живые организмы, пока видятся нам как нечто фантастическое, нереальное. Но то, что нереально сегодня, уже завтра может оказаться чем-то обыденным и настолько естественным, что трудно будет представить, как без этого можно было обходиться в прошлом.
Итак, ДНК-вычисления - это раздел области молекулярных вычислений на границе молекулярной биологии и компьютерных наук. Основная идея ДНК-вычислений - построение новой парадигмы, создание новых алгоритмов вычислений на основе знаний о строении и функциях молекулы ДНК и операций, которые выполняются в живых клетках над молекулами ДНК при помощи различных ферментов. К перспективам ДНК-вычислений относится создание биологического нанокомпьютера, который будет способен хранить терабайты информации при объеме в несколько микрометров. Такой компьютер можно будет вживлять в клетку живого организма, а его производительность будет исчисляться миллиардами операций в секунду при энергопотреблении не более одной миллиардной доли ватта.
Преимущества ДНК в компьютерных технологиях
Для современных процессоров и микросхем в качестве строительного материала используется кремний. Но возможности кремния не беспредельны, и в конечном счете мы подойдем к той черте, когда дальнейший рост вычислительной мощности процессоров окажется исчерпан. А потому перед человечеством уже сейчас остро стоит проблема поиска новых технологий и материалов, которые смогли бы в будущем заменить кремний.
Молекулы ДНК могут оказаться тем самым материалом, который впоследствии заменит кремниевые транзисторы с их бинарной логикой. Достаточно сказать, что всего один фунт (453 г) ДНК-молекул обладает емкостью для хранения данных, которая превосходит суммарную емкость всех современных электронных систем хранения данных, а вычислительная мощность ДНК-процессора размером с каплю будет выше самого мощного современного суперкомпьютера.
Более 10 триллионов ДНК-молекул занимают объем всего в 1 см3. Однако такого количества молекул достаточно для хранения объема информации в 10 Тбайт, при этом они могут производить 10 трлн операций в секунду.
Еще одно преимущество ДНК-процессоров в сравнении с обычными кремниевыми процессорами заключается в том, что они могут производить все вычисления не последовательно, а параллельно, что обеспечивает выполнение сложнейших математических расчетов буквально за считаные минуты. Традиционным компьютерам для выполнения таких расчетов потребовались бы месяцы и годы.
Строение молекул ДНК
Как известно, современные компьютеры работают с бинарной логикой, подразумевающей наличие всего двух состояний: логического нуля и единицы. Используя двоичный код, то есть последовательность нулей и единиц, можно закодировать любую информацию. В молекулах ДНК имеется четыре базовых основания: аденин (A), гуанин (G), цитозин (C) и тимин (T), связанных друг с другом в цепочку. То есть молекула ДНК (одинарная цепочка) может иметь, например, такой вид: ATTTACGGCC - здесь используется не двоичная, а четверичная логика. И подобно тому, как в двоичной логике любую информацию можно закодировать в виде последовательности нулей и единиц, в молекулах ДНК можно кодировать любую информацию путем сочетания базовых оснований.
Базовые основания в молекулах ДНК находятся друг от друга на расстоянии 0,34 нанометра, что обусловливает их огромную информативную емкость - линейная плотность составляет 18 Мбит/дюйм. Если же говорить о поверхностной информативной плотности, предполагая, что на одно базовое основание приходится площадь в 1 квадратный нанометр, то она составляет более миллиона гигабит на квадратный дюйм. Для сравнения отметим, что поверхностная плотность записи современных жестких дисков составляет порядка 7 Гбит/дюйм 2.
Другое важное свойство ДНК-молекул заключается в том, что они могут иметь форму регулярной двойной спирали, диаметр которой составляет всего 2 нм. Такая спираль состоит из двух цепей (последовательностей базовых оснований), причем содержание первой цепи строго соответствует содержанию второй.
Это соответствие достигается благодаря наличию водородных связей между направленными навстречу друг другу основаниями двух цепей - попарно G и C или A и T. Описывая это свойство двойной спирали, молекулярные биологи говорят, что цепи ДНК комплементарны за счет образования пар G-C и A-T.
К примеру, если последовательность S записывается как ATTACGTCG, то дополняющая ее последовательность S’ будет иметь вид TAATGCAGC.
Процесс соединения двух одинарных цепочек ДНК путем связывания комплементарных оснований в регулярную двойную спираль называется ренатурацией, а обратный процесс, то есть разъединение двойной цепочки и получение двух одинарных цепочек, - денатурацией (рис. 1).
Рис. 1. Процессы ренатурации и денатурации
Комплементарная особенность строения ДНК-молекул может использоваться при ДНК-вычислениях. К примеру, на основе дополняющих друг друга последовательностей можно реализовать мощнейший механизм коррекции ошибок, который чем-то напоминает технологию зеркалирования данных RAID Level 1.
Базовые операции над ДНК-молекулами
Для различных манипуляций над ДНК-молекулами используются различные энзимы (ферменты). И точно так же, как современные микропроцессоры имеют набор базовых операций типа сложения, сдвига, логических операций AND, OR и NOT NOR, ДНК-молекулы под воздействием энзимов могут выполнять такие базовые операции, как разрезание, копирование, вставка и др. Причем все операции над ДНК-молекулами можно производить параллельно и независимо от других операций, к примеру дополнение цепочки ДНК осуществляется при воздействии на исходную молекулу ферментов - полимераз. Для работы полимеразы необходимо наличие одноцепочечной молекулы (матрицы), определяющей добавляемую цепочку по принципу комплементарности, праймера (небольшого двухцепочечного участка) и свободных нуклеотидов в растворе. Процесс дополнения цепочки ДНК показан на рис. 2.
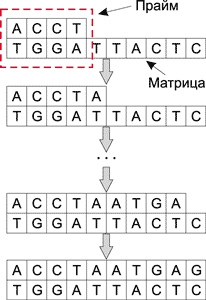
Рис. 2. Процесс дополнения цепочки ДНК
при воздействии на исходную молекулу полимеразы
Существуют полимеразы, которым не требуются матрицы для удлинения цепочки ДНК. Например, терминальная трансфераза добавляет одинарные цепочки ДНК к обоим концам двухцепочечной молекулы. Таким образом можно конструировать произвольную цепь ДНК (рис. 3).

Рис. 3. Процесс удлинения цепочки ДНК
За укорачивание и разрезание молекул ДНК отвечают ферменты - нуклеазы. Различают эндонуклеазы и экзонуклеазы. Последние могут укорачивать и одноцепочечные и двухцепочечные молекулы с одного или с обоих концов (рис. 4), а эндонуклеазы - только с концов.

Рис. 4. Процесс укорачивания молекулы
ДНК под воздействием экзонуклеазы
Разрезание молекул ДНК возможно под воздействием сайт-специфичных эндонуклеазов - рестриктазов, которые разрезают их в определенном месте, закодированном последовательностью нуклеотидов (сайтом узнавания). Разрез может быть прямым или несимметричным и проходить по сайту узнавания либо вне его. Эндонуклеазы разрушают внутренние связи в молекуле ДНК (рис. 5).

Рис. 5. Разрезание молекулы ДНК
под воздействием рестриктазов
Сшивка - операция, обратная разрезанию, - происходит под воздействием ферментов - лигазов. «Липкие концы» соединяются вместе с образованием водородных связей. Лигазы служат для того, чтобы закрыть насечки, то есть способствовать образованию в нужных местах фосфодиэфирных связей, соединяющих основания друг с другом в пределах одной цепочки (рис. 6).

Рис. 6. Сшивка ДНК-молекул под воздействием лигазов
Еще одна интересная операция над ДНК-молекулами, которую можно отнести к числу базовых, - это модификация. Она используется для того, чтобы рестриктазы не смогли найти определенный сайт и не разрушили молекулу. Существует несколько типов модифицирующих ферментов - метилазы, фосфатазы и т.д.
Метилаза имеет тот же сайт узнавания, что и соответствующая рестриктаза. При нахождении нужной молекулы метилаза модифицирует участок с сайтом так, что рестриктаза уже не сможет идентифицировать эту молекулу.
Копирование, или размножение, ДНК-молекул осуществляется в ходе полимеразной цепной реакции (Polymerase Chain Reaction, PCR) - рис. 7. Процесс копирования можно разделить на несколько стадий: денатурация, праймирование и удлинение. Он происходит лавинообразно. На первом шаге из одной молекулы образуются две, на втором - из двух молекул - четыре, а после n-шагов получается уже 2n молекул.

Рис. 7. Процесс копирования ДНК-молекулы
Еще одна операция, которую можно производить над ДНК-молекулами, - это секвенирование, то есть определение последовательности нуклеотидов в ДНК. Для секвенирования цепочек разной длины применяют различные методы. При помощи метода праймер-опосредованной прогулки удается на одном шаге секвенировать последовательность в 250-350 нуклеотидов. После открытия рестриктаз стало возможным секвенировать длинные последовательности по частям.
Ну и последняя процедура, которую мы упомянем, - это гель-электрофорез, используемый для разделения молекул ДНК по длине. Если молекулы поместить в гель и приложить постоянное электрическое поле, то они будут двигаться по направлению к аноду, причем более короткие молекулы будут двигаться быстрее. Используя данное явление, можно реализовать сортировку ДНК-молекул по длине.
ДНК-вычисления
ДНК-молекулы со своей уникальной формой строения и возможностью реализовать параллельные вычисления позволяют по-другому взглянуть на проблему компьютерных вычислений. Традиционные процессоры выполняют программы последовательно. Несмотря на существование многопроцессорных систем, многоядерных процессоров и различных технологий, направленных на повышение уровня параллелизма, в своей основе все компьютеры, построенные на основе фон-неймановской архитектуры, являются устройствами с последовательным режимом выполнения команд. Все современные процессоры реализуют следующий алгоритм обработки команд и данных: выборка команд и данных из памяти и исполнение инструкций над выбранными данными. Этот цикл повторяется многократно и с огромной скоростью.
ДНК-вычисления имеют в своей основе абсолютно иную, параллельную архитектуру и в ряде случаев именно благодаря этому способны с легкостью рассчитывать те задачи, для решения которых компьютерам на базе фон-неймановской архитектуры потребовались бы годы.
Эксперимент Эдлмана
История ДНК-вычислений начинается в 1994 году. Именно тогда Леонард М. Эдлман (Leonard M. Adleman) попытался решить весьма тривиальную математическую задачу абсолютно нетривиальным способом - с использованием ДНК-вычислений. Фактически это было первой демонстраций прототипа биологического компьютера на основе ДНК-вычислений.
Задача, которую Эдлман выбрал для выполнения с помощью ДНК-вычислений, известна как поиск гамильтонова пути в графе или выбор маршрута путешествия (traveling salesman problem). Смысл ее заключается в следующем: имеется несколько городов, которые необходимо посетить, причем побывать в каждом городе можно только один раз.
Зная пункт отправления и конечный пункт, необходимо определить маршрут путешествия (если он существует). При этом маршрут составляется с учетом возможных авиаперелетов и коннектов различных авиарейсов.
Итак, предположим, что имеется всего четыре города (в эксперименте Эдлмана использовалось семь городов): Атланта (Atlanta), Бостон (Boston), Детройт (Detroit) и Чикаго (Chicago). Перед путешественником ставится задача выбрать маршрут, чтобы попасть из Атланты в Детройт, побывав при этом в каждом городе только один раз. Схемы возможных сообщений между городами показаны на рис. 8.

Рис. 8. Схемы возможных сообщений
между городами
Нетрудно заметить (для этого требуется всего несколько секунд), что единственно возможный маршрут (гамильтонов путь) следующий: Атланта - Бостон - Чикаго - Детройт.
Действительно, при небольшом количестве городов составить такой маршрут довольно просто. Но с увеличением их числа сложность решения задачи экспоненциально возрастает и становится трудновыполнимой не только для человека, но и для компьютера.
Так, на рис. 9 показан граф из семи вершин с указанием возможных переходов между ними. Для поиска гамильтонова пути обычному человеку требуется не более одной минуты. Именно такой граф был использован в эксперименте Эдлмана. На рис. 10 представлен граф из 12 вершин - в этом случае поиск гамильтонова пути оказывается уже не такой простой задачей. Вообще, сложность решения задачи поиска гамильтонова пути возрастает экспоненциально с ростом числа вершин в графе. К примеру, для графа из 10 вершин существует 106 возможных путей; для графа из 20 вершин - 1012, а для графа из 100 вершин - 10100 вариантов. Понятно, что в последнем случае для генерации всех возможных путей и их проверки потребуется огромное время даже для современного суперкомпьютера.

Рис. 9. Поиск оптимального маршрута путешествия

Рис. 10. Граф, состоящий из 12 вершин
Итак, вернемся к нашему примеру с поиском гамильтонова пути в случае четырех городов (см. рис. 8).
Для решения данной задачи с использованием ДНК-вычислений Эдлман закодировал название каждого города в виде одной цепочки ДНК, причем каждая из них содержала 20 базовых оснований. Для простоты мы будем кодировать каждый город ДНК-цепочкой из восьми оснований. ДНК-коды городов показаны в табл. 1. Обратите внимание, что цепочка длиной в восемь базовых оснований оказывается избыточной для кодирования всего четырех городов.

Таблица 1. ДНК-коды городов
Отметим, что для каждого ДНК-кода города, который определяет одинарную ДНК-цепочку, существует и комплементарная цепочка, то есть комплементарный ДНК-код города, причем и ДНК-код города, и комплементраный код абсолютно равноправны.
Далее с помощью одинарных ДНК-цепочек необходимо закодировать все возможные перелеты (Атланта - Бостон, Бостон - Детройт, Чикаго - Детройт и т.д.). Для этого использовался следующий подход. Из названия города отправления брались четыре последних базовых основания, а из названия города прибытия - четыре первых.
К примеру, перелету Атланта - Бостон будет соответствовать следующая последовательность: GCAG TCGG (рис. 11).

Рис. 11. Кодирование перелетов между городами
ДНК-кодирование всех возможных перелетов показано в табл. 2.

Таблица 2. ДНК-коды всех возможных перелетов
Итак, после того как готовы коды городов и возможных перелетов между ними, можно непосредственно переходить к вычислению гамильтонова пути. Процесс вычисления состоит из четырех этапов:
- Сгенерировать все возможные маршруты.
- Отобрать маршруты, которые начинаются в Атланте и заканчиваются Детройтом.
- Выбрать маршруты, длина которых соответствует количеству городов (в нашем случае длина маршрута составляет четыре города).
- Выбрать маршруты, в которых каждый город присутствует только один раз.
Итак, на первом этапе мы должны сгенерировать все возможные маршруты. Напомним, что правильный маршрут соответствует перелетам Атланта - Бостон - Чикаго - Детройт. Этому маршруту соответствует ДНК-молекула GCAG TCGG ACTG GGCT ATGT CCGA.
Для того чтобы сгенерировать все возможные маршруты достаточно поместить в пробирку все необходимые и заранее заготовленные ингредиенты, то есть ДНК-молекулы, соответствующие всем возможным перелетам, и ДНК-молекулы, соответствующие всем городам. Но вместо того, чтобы применять одинарные ДНК-цепочки, соответствующие названиям городов, необходимо использовать комплементарные им ДНК-цепочки, то есть вместо ДНК-цепочки ACTT GCAG, соответствующей Атланте, будем применять комплементарную ДНК-цепочку TGAA CGTC и т.д., поскольку ДНК-код города и комплементраный код абсолютно равноправны.
Далее все эти молекулы (достаточно буквально щепотки, которая будет содержать порядка 1014 различных молекул) помещаем в воду, добавляем лигазов, произносим заклинание и… буквально через несколько секунд получаем все возможные маршруты.
Процесс образования цепочек ДНК, соответствующих различным маршрутам, происходит следующим образом. Рассмотрим, к примеру, цепочку GCAG TCGG, отвечающую за перелет Атланта - Бостон. Вследствие высокой концентрации различных молекул, данная цепочка обязательно встретится с комплементарной ДНК-цепочкой AGCC TGAC, соответствующей Бостону. Поскольку группы TCGG и AGCC комплементарны друг другу, то за счет образования водородных связей эти цепочки сцепятся друг с другом (рис. 12).
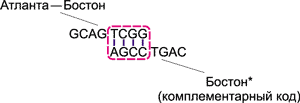
Рис. 12. Сцепление цепочек, соответствующих
перелету Атланта - Бостон и Бостону
Теперь образовавшаяся цепочка неминуемо встретится с ДНК-цепочкой ACTG GGCT, соответствующей авиаперелету Бостон - Чикаго, и поскольку группа ACTG (первые четыре основания в этой цепочке) комплементарна группе TGAC (последние четыре основания в комплементарном коде Бостона), то ДНК-цепочка ACTG GGCT присоединится к уже образовавшейся цепочке. Далее к этой цепочке таким же образом присоединится ДНК-цепочка, соответствующая городу Чикаго (комплементарный код), а затем и цепочка авиаперелета Чикаго - Детройт. Процесс образования маршрута показан на рис. 13.

Рис. 13. Процесс образования ДНК-цепочки, соответствующей маршруту
Атланта - Бостон - Чикаго - Детройт
Мы рассмотрели пример образования только одного маршрута (причем это именно гамильтонов маршрут). Аналогичным образом получаются и все остальные возможные маршруты (например, Атланта - Бостон - Атланта - Детройт). Важно, что все маршруты формируются одновременно, то есть параллельно. Причем время, требуемое для создания всех возможных маршрутов в данной задаче и всех маршрутов в задаче с 10 или 20 городами, абсолютно одинаково (лишь бы хватило исходных ДНК-молекул). Собственно, именно в параллельном алгоритме ДНК-вычислений и заключается основное преимущество в сравнении с фон-неймановской архитектурой.
Итак, в пробирке образованы ДНК-молекулы, соответствующие всем возможным маршрутам. Однако это еще не решение задачи - нам необходимо выделить ту единственную ДНК-молекулу, которая отвечает за гамильтонов маршрут. Поэтому на следующем этапе необходимо отобрать молекулы, соответствующие маршрутам, начинающимся в Атланте и заканчивающимся в Детройте.
Для этого используется полимеразная цепная реакция (PCR), в результате которой создается множество копий только тех ДНК-цепочек, которые начинаются с кода Атланты и заканчиваются кодом Детройта.
Для реализации полимеразной цепной реакции применяются два прайма: GCAG и GGCT. Процесс копирования ДНК-модекул, начинающихся с ДНК-кода Атланты и заканчивающихся ДНК-кодом Детройта, показан на рис. 14.

Рис. 14. Процесс копирования ДНК-молекул в ходе PCR-реакции
Отметим, что в присутствии праймов GCAG и GGCT будут копироваться и те ДНК-молекулы, которые начинаются с ДНК-кодов Атланты, но не заканчиваются ДНК-кодом Детройта (под действием прайма GCAG), а также ДНК-молекулы, которые заканчиваются ДНК-кодом Детройта, но не начинаются с ДНК-кода Атланты (под действием прайма GGCT). Понятно, что скорость копирования таких молекул будет гораздо ниже скорости копирования ДНК-молекул, начинающихся с ДНК-кода Атланты и заканчивающихся ДНК-кодом Детройта. Следовательно, после PCR-реакции мы получим преобладающее количество ДНК-молекул в форме регулярной двойной спирали, соответствующих маршрутам, начинающимся в Атланте и заканчивающимся в Детройте.
На следующем этапе необходимо выделить молекулы нужной длины, то есть те, что содержат ДНК-коды ровно четырех городов. Для этого используется гель-электрофорез, что позволяет отсортировать молекулы по длине. В результате мы получаем молекулы нужной длины (ровно четыре города), начинающиеся с кода Атланты и заканчивающиеся кодом Детройта.
Теперь необходимо убедиться, что в отобранных молекулах код каждого города присутствует только один раз. Эта операция реализуется с применением процесса, известного как affinity purification.
Для данной операции используется микроскопический магнитный шарик диаметром порядка одного микрона. К нему притягиваются комлементарные ДНК-коды того или иного города, которые выполняют функцию пробы. К примеру, если требуется проверить, присутствует ли в исследуемой ДНК-цепочке код города Бостона, то необходимо сначала поместить магнитный шарик в пробирку с ДНК-молекулами, соответствующими ДНК-кодам Бостона. В результате мы получим магнитный шарик, облепленный нужными нам пробами. Затем этот шарик помещается в пробирку с исследуемыми ДНК-цепочками - в результате к нему (за счет образования водородных связей между комплементарными группами) притянутся ДНК-цепочки, в которых присутствует комплементарный код Бостона. Далее шарик с отсортированными молекулами вынимается и помещается в новый раствор, из которого затем удаляется (при повышении температуры ДНК-молекулы отваливаются от шарика). Данная процедура (сортировка) повторяется последовательно для каждого города, и в результате мы получаем только те молекулы, в которых содержатся ДНК-коды всех городов, а значит, и маршруты, соответствующие гамильтонову пути. Фактически задача решена - осталось лишь просчитать ответ.
Заключение
Эдлман продемонстрировал решение задачи поиска гамильтонова пути на примере всего семи городов и потратил на это семь дней. Это был первый эксперимент, продемонстрировавший возможности ДНК-вычислений. Фактически Эдлман доказал, что, пользуясь вычислениями на ДНК, можно эффективно решать задачи переборного характера, и обозначил технику, которая в дальнейшем послужила основой для создания модели параллельной фильтрации.
Впрочем, многие исследователи не испытывают оптимизма по поводу будущего биологических компьютеров. Вот лишь маленький пример. Если бы подобным методом понадобилось найти гамильтонов путь в графе, состоящем из 200 вершин, потребовалось бы количество ДНК-молекул, сопоставимое по весу со всей нашей планетой! Это принципиальное ограничение, конечно же, является своего рода тупиковой ситуацией. Поэтому многие исследовательские лаборатории (например, компания IBM) предпочли сфокусировать свое внимание на других идеях альтернативных компьютеров, таких как углеродные нанотрубки и квантовые компьютеры.
После эксперимента Эдлмана было проведено множество других исследований возможностей ДНК-вычислений. Например, можно вспомнить опыт Э.Шапиро: в нем был реализован конечный автомат, который может находиться в двух состояниях: S0 и S1 - и отвечает на вопрос: четное или нечетное количество символов содержится во входной последовательности символов.
Сегодня ДНК-вычисления - это не более чем перспективные технологии на уровне лабораторных исследований, причем в таком состоянии они будут находиться еще не один год. Фактически на современном этапе развития необходимо ответить на следующий глобальный вопрос: какой класс задач поддается решению при помощи ДНК и можно ли построить общую модель ДНК-вычислений, пригодную как для реализации, так и для использования?
Мономерными звеньями которого являются нуклиатиды.
Что такое ДНК?
Вся информация о строении и функционировании любого живого организма содержится в закодированном виде в его генетическом материале. Основу генетического материала организма составляет дезоксирибонуклеиновая кислота (ДНК) .
ДНК большинства организмов – это длинная двухцепочечная полимерная молекула. Последовательность мономерных звеньев (дезоксирибонуклеотидов ) в одной ее цепи соответствует (комплементарна ) последовательности дезоксирибонуклеотидов в другой. Принцип комплементарности обеспечивает синтез новых молекул ДНК, идентичных исходным, при их удвоении (репликации ).
Участок молекулы ДНК, кодирующий определенный признак, – ген .
Гены – это индивидуальные генетические элементы, имеющие строго специфичную нуклеотидную последовательность, и кодирующие определенные признаки организма. Одни из них кодируют белки, другие - только молекулы РНК.
Информация, которая содержится в генах, кодирующих белки (структурных генах), расшифровывается в ходе двух последовательных процессов:
- синтеза РНК (транскрипции ): на определенном участке ДНК как на матрице синтезируется матричная РНК (мРНК).
- синтеза белка (трансляции): В ходе согласованной работы многокомпонентной системы при участии транспортных РНК (тРНК ), мРНК , ферментов и различных белковых факторов осуществляется синтез белковой молекулы .
Все эти процессы обеспечивают правильный перевод зашифрованной в ДНК генетической информации с языка нуклеотидов на язык аминокислот. Аминокислотная последовательность белковой молекулы определяет ее структуру и функции.
Строение ДНК
ДНК – это линейный органический полимер . Его – нуклеотиды , которые, в свою очередь, состоят из:
При этом, фосфатная группа присоединена к 5′-атому углерода моносахаридного остатка, а органическое основание - к 1′-атому .
Основания в ДНК бывают двух типов:

Строение нуклеотидов в молекуле ДНК
В ДНК моносахарид представлен 2′-дезоксирибозой , содержащей только 1 гидроксильную группу (ОН) , а в РНК - рибозой , имеющей 2 гидроксильные группы (OH ).
Нуклеотиды соединены друг с другом фосфодиэфирными связями , при этом фосфатная группа 5′-углеродного атома одного нуклеотида связана с З’-ОН-группой дезоксирибозы соседнего нуклеотида (рисунок 1). На одном конце полинуклеотидной цепи находится З’-ОН-группа (З’-конец), а на другом - 5′-фосфатная группа (5′-конец).
Уровни структуры ДНК
Принято выделять 3 уровня структуры ДНК:
- первичную;
- вторичную;
- третичную.
Первичная структура ДНК – это последовательность расположения нуклеотидов в полинуклеотидной цепи ДНК.
Вторичная структура ДНК стабилизируется между комплементарными парами оснований и представляет собой двойную спираль из двух антипараллелных цепочек, закрученных вправо вокруг одной оси.
Общий виток спирали- 3,4нм , расстояние между цепочками 2нм.
Третичная структура ДНК – суперсперализация ДНК. Двойная спираль ДНК на некоторых участках может подвергаться дальнейшей спирализации с образованием суперспирали или открытой кольцевой формы, что часто вызвано ковалентным соединением их открытых концов. Суперспиральная структура ДНК обеспечивает экономную упаковку очень длинной молекулы ДНК в хромосоме. Так, в вытянутой форме длина молекулы ДНК составляет 8 см , а в форме суперспирали укладывается в 5 нм .
Правило Чаргаффа
Правило Э. Чаргаффа – это закономерность количественного содержания азотистых оснований в молекуле ДНК:
- У ДНК молярные доли пуриновых и пиримидиновых оснований равны: А+ G = C + Т или (А + G )/(C + Т)=1 .
- В ДНК количество оснований с аминогруппами (А + C ) равно количеству оснований с кетогруппами (G + Т): А + C = G + Т или (А + C )/(G + Т)= 1
- Правило эквивалентности, то есть: А=Т, Г=Ц; А/Т = 1; Г/Ц=1.
- Нуклеотидный состав ДНК у организмов различных групп специфичен и характеризуется коэффициентом специфичности: (Г+Ц)/(А+Т). У высших растений и животных коэффициент специфичности меньше 1, и колеблется незначительно: от 0,54 до 0,98 , у микроорганизмов он больше 1.
Модель ДНК Уотсона-Крика
Б 1953 г. Джеймс Уотсон и Фрэнсис Крик , основываясь на данных рентгеноструктурного анализа кристаллов ДНК, пришли к выводу, что нативная ДНК состоит из двух полимерных цепей, образующих двойную спираль (рисунок 3).
Навитые одна на другую полинуклеотидные цепи удерживаются вместе водородными связями , образующимися между комплементарными основаниями противоположных цепей (рисунок 3). При этом аденин образует пару только с тимином , а гуанин - с цитозином . Пара оснований А-Т стабилизируется двумя водородными связями , а пара G-С - тремя .
Длина двухцепочечной ДНК обычно измеряется числом пар комплементарных нуклеотидов (п .н .). Для молекул ДНК, состоящих из тысяч или миллионов пар нуклеотидов, приняты единицы т.п.н. и м.п.н. соответственно. Например, ДНК хромосомы 1 человека представляет собой одну двойную спираль длиной 263 м.п.н .
Сахарофосфатный остов молекулы , который состоит из фосфатных групп и дезоксирибозных остатков, соединенных 5’-З’-фосфодиэфирными связями , образует «боковины винтовой лестницы», а пары оснований А-Т и G-С - ее ступеньки (рисунок 3).
Рисунок 3: Модель ДНК Уотсона-Крика
Цепи молекулы ДНК антипараллельны : одна из них имеет направление 3’→5′ , другая 5’→3′ . В соответствии с принципом комплементарности , если в одной из цепей имеется нуклеотидная последовательность 5-TAGGCAT-3′ , то в комплементарной цепи в этом месте должна находиться последовательность 3′-ATCCGTA-5′ . В этом случае двухцепочечная форма будет выглядеть следующим образом:
- 5′-TAGGCAT-3′
- 3-ATCCGTA-5′.
В такой записи 5′-конец верхней цепи всегда располагают слева, а 3′-конец - справа.
Носитель генетической информации должен удовлетворять двум основным требованиям: воспроизводиться (реплицироваться) с высокой точностью и детерминировать (кодировать) синтез белковых молекул .
Модель ДНК Уотсона-Крика полностью отвечает этим требованиям, так как:
- согласно принципу комплементарности каждая цепь ДНК может служить матрицей для образования новой комплементарной цепи. Следовательно, после одного раунда образуются две дочерние молекулы, каждая из которых имеет такую же нуклеотидную последовательность, как исходная молекула ДНК.
- нуклеотидная последовательность структурного гена однозначно задает аминокислотную последовательность кодируемого ею белка.
- Одна молекула ДНК человека вмещает порядка 1,5 гигабайта информации . При этом, ДНК всех клеток человеческого организма занимают 60 млрд. терабайт, что сохраняются на 150-160 граммах ДНК.
- Международный день ДНК отмечается 25 апреля. Именно в этот день в 1953 году Джеймс Уотсон и Фрэнсис Крик опубликовали в журнале Nature свою статью под названием «Молекулярная структура нуклеиновых кислот» , где описали двойную спираль молекулы ДНК.
Список литературы: Молекулярная биотехнология: принципы и применение, Б.Глик, Дж. Пастернак, 2002 год
ДНК (дезоксирибонуклеиновая кислота) – биологический полимер, состоящий из двух полинуклеотидных цепей, соединенных друг с другом. Мономеры, составляющие каждую из цепей ДНК, представляют собой сложные органические соединения, включающие одно из четырех азотистых оснований: аденин (А) или тимин (Т), цитозин (Ц) или гуанин (Г), пятиатомный сахар пентозу – дезоксирибозу, по имени которой получила название и сама ДНК, а также остаток фосфорной кислоты. Эти соединения носят название нуклеотидов.
Эти цепи соединяются друг с другом водородными связями между их азотистыми основаниями по принципу комплементарности. Аденин одной цепи соединяется двумя водородными связями с тимином другой цепи, а между гуанином и цитозином разных цепей образуются три водородные связи. Такое соединение азотистых оснований обеспечивает прочную связь двух цепей и сохранение равного расстояния между ними на всем протяжении.
Другой важной особенностью объединения двух полинуклеотидных цепей в молекуле ДНК является их антипараллельность: 5"-конец одной цепи соединяется с 3"-концом другой, и наоборот.
Молекула ДНК, сост. Из двух цепей, образует спираль, закрученную вокруг собственной оси. Диаметр спирали составляет 2 нм, длина шага - 3, 4 нм. В каждый виток входит 10 пар нуклеотидов.
* Чаще всего двойные спирали являются правозакрученными. Большинство молекул ДНК в растворе находится в правозакрученной - В-форме (В-ДНК). Однако встречаются также левозакрученные формы (Z-ДНК). Какое количество этой ДНК присутствует в клетках и каково ее биологическое значение, пока не установлено.
* Таким образом, в структурной организации молекулы ДНК можно выделить первичную структуру - полинуклеотидную цепь, вторичную структуру - две комплементарные друг другу и антипараллельные полинуклеотидные цепи, соединенные водородными связями, и третичную структуру - трехмерную спираль с приведенными выше пространственными характеристиками.
9. Типы рнк в клетке. Функции различных рнк
Роль посредника, функцией которого является перевод наследственной информации, сохраняемой в ДНК, в рабочую форму, играют рибонуклеиновые кислоты - РНК.
Известны двух – и одно цепочечные молекулы РНК. Двухцепочечные РНК служат для хранения и воспроизведения наследственной информации у некоторых вирусов, т.е. у них выполняется функции хромосом. Одноцепочечные РНК осуществляют перенос информации о последовательности аминокислот в белках от хромосомы к месту их синтеза и участвуют в процессах синтеза.
В отличие от молекул ДНК рибонуклеиновые кислоты представлены одной полинуклеотидной цепью, которая состоит из четырех разновидностей нуклеотидов, содержащих сахар, рибозу, фосфат и одно из четырех азотистых оснований - аденин, гуанин, урацил или цитозин. РНК синтезируется на молекулах ДНК при помощи ферментов РНК-полимераз с соблюдением принципа комплементарности и антипараллельности, причем аденину ДНК в РНК комплементарен урацил. Все многообразие РНК, действующих в клетке, можно разделить на три основных вида: мРНК, тРНК, рРНК.
Матричная, или информационная, РНК (мРНК, или иРНК). Транскрипция. Для того чтобы синтезировать белки с заданными свойствами, к месту их построения поступает «инструкция» о порядке включения аминокислот в пептидную цепь. Эта инструкция заключена в нуклеотидной последовательности матричных, или информационных РНК (мРНК, иРНК), синтезируемых на соответствующих участках ДНК. Процесс синтеза мРНК называют транскрипцией.
Синтез мРНК начинается с обнаружения РНК-полимеразой особого участка в молекуле ДНК, который указывает место начала транскрипции - промотора. После присоединения к промотору РНК-полимераза раскручивает прилежащий виток спирали ДНК. Две цепи ДНК в этом месте расходятся, и на одной из них фермент осуществляет синтез мРНК. Сборка рибонуклеотидов в цепь происходит с соблюдением их комплементарности нуклеотидам ДНК, а также антипараллельно по отношению к матричной цепи ДНК. В связи с тем, что РНК-полимераза способна собирать полинуклеотид лишь от 5"-конца к 3"-концу, матрицей для транскрипции может служить только одна из двух цепей ДНК, а именно та, которая обращена к ферменту своим 3"-концом (3" → 5"). Такуюцепь называют кодогенной
тРНК - РНК, функцией которой является транспортировка аминокислот к месту синтеза белка. тРНК также принимают непосредственное участие в наращивании полипептидной цепи, присоединяясь - будучи в комплексе с аминокислотой - к кодону мРНК и обеспечивая необходимую для образования новой пептидной связи конформацию комплекса.
Для каждой аминокислоты существует своя тРНК.тРНК является одноцепочечной РНК, однако в функциональной форме имеет конформацию «листа клевера» или «кловерлиф» (англ. cloverleaf). Аминокислота ковалентно присоединяется к 3"-концу молекулы с помощью специфичного для каждого типа тРНК фермента аминоацил-тРНК-синтетазы. На участке C находится антикодон, соответствующий аминокислоте.
(рРНК) - несколько молекул РНК, составляющих основу рибосомы. Основной функцией рРНК является осуществление процесса трансляции - считывания информации с мРНК при помощи адапторных молекул тРНК и катализ образования пептидных связей между присоединёнными к тРНК аминокислотами.
Рибосомные РНК являются не только структурным компонентом рибосом, но и обеспечивают связывание их с определенной нуклеотидной последовательностью мРНК. Этим устанавливаются начало и рамка считывания при образовании пептидной цепи. Кроме того, они обеспечивают взаимодействие рибосомы и тРНК. Многочисленные белки, входящие в состав рибосом наряду с рРНК, выполняют как структурную, так и ферментативную роль.
Многих людей всегда интересовало, почему некоторые признаки, имеющиеся у родителей, передаются ребенку (например, цвет глаз, волос, форма лица и другие). Наукой было доказано, что данная передача признака зависит от генетического материала, или ДНК.
Что такое ДНК?
Нуклеотид
Как было сказано, основной структурной единицей дезоксирибонуклеиновой кислоты является нуклеотид. Это сложное образование. Состав нуклеотида ДНК следующий.
По центру нуклеотида находится пятикомпонентный сахар (в ДНК в отличие от РНК, в которой содержится рибоза). К нему присоединяется азотистое основание, которых выделяют 5 типов: аденин, гуанин, тимин, урацил и цитозин. Кроме того, каждый нуклеотид имеет в своем составе и остаток фосфорной кислоты.

В состав ДНК входят только те нуклеотиды, которые имеют указанные структурные единицы.
Все нуклеотиды расположены в виде цепи и следуют друг за другом. Группируясь по триплетам (по три нуклеотида), они образуют последовательность, в которой каждый триплет соответствует определенной аминокислоте. В результате образуется цепь.
Они объединяются между собой за счет связей азотистых оснований. Основная связь между нуклеотидами параллельных цепей - водородная.
Нуклеотидные последовательности являются основой генов. Нарушение в их структуре ведет к сбою в синтезе белков и проявлению мутаций. В состав ДНК входят одинаковые гены, определяющиеся практически у всех людей и отличающие их от других организмов.
Модификация нуклеотида
В некоторых случаях для более стабильной передачи того или иного признака используется модифицирование азотистого основания. Химический состав ДНК изменяется за счет присоединения метильной группы (СН3). Подобная модификация (на одном нуклеотиде) позволяет стабилизировать генную экспрессию и передачу признаков дочерним клеткам.

Подобное “улучшение” структуры молекулы никоим образом не сказывается на объединении азотистых оснований.
Данная модификация используется и при инактивации Х-хромосомы. В результате этого образуются тельца Барра.
При усиленном канцерогенезе анализ ДНК показывает, что цепочка нуклеотидов была подвержена метилированию на многих основаниях. В проведенных наблюдениях было замечено, что источником мутации обычно служит метилированный цитозин. Обычно при опухолевом процессе деметилирование может способствовать остановке процесса, но за счет своей сложности данная реакция не проводится.
Структура ДНК
В строении молекулы выделяют два типа структуры. Первый тип - линейная последовательность, образованная нуклеотидами. Их построение подчиняется некоторым законам. Запись нуклеотидов на молекуле ДНК начинается с 5’-конца и заканчивается 3’-концом. Вторая цепь, расположенная напротив, строится таким же образом, только в пространственном отношении молекулы находятся одна напротив другой, причем 5’-конец одной цепи расположен напротив 3’-конца второй.

Вторичная структура ДНК - спираль. Обуславливается наличием водородных связей между располагающимися друг напротив друга нуклеотидами. Водородная связь образуется между комплементарными азотистыми основаниями (например, напротив аденина первой цепи может находиться только тимин, а напротив гуанина - цитозин либо урацил). Подобная точность обусловлена тем, что построение второй цепи происходит на основе первой, поэтому между азотистыми основаниями наблюдается точное соответствие.
Синтез молекулы
Каким же образом образуется молекула ДНК?
В цикле ее образования выделяют три стадии:
- Рассоединение цепей.
- Присоединение синтезирующих единиц к одной из цепей.
- Достраивание второй цепи по принципу комплементарности.
На стадии разъединения молекулы основную роль играют ферменты - ДНК-гиразы. Данные ферменты ориентированы на разрушение водородных связей между цепями.

После расхождения цепей в дело вступает основной синтезирующий фермент - ДНК-полимераза. Ее присоединение наблюдается на участке 5’. Далее данный фермент движется в сторону 3’-конца, попутно присоединяя необходимые нуклеотиды с соответствующими азотистыми основаниями. Дойдя до определенного участка (терминатора) на 3’-конце, полимераза отсоединяется от исходной цепи.
После того как образовалась дочерняя цепь, между основаниями образуется водородная связь, которая и скрепляет вновь образованную молекулу ДНК.
Где можно найти данную молекулу?
Если углубиться в строение клеток и тканей, то можно увидеть, что ДНК в основном содержится в отвечает за образование новых, дочерних, клеток или их клонов. При этом находящаяся в нем, разделяется между новообразованными клетками равномерно (образуются клоны) или по частям (часто можно наблюдать такое явление при мейозе). Поражение ядра влечет за собой нарушение образования новых тканей, что приводит к мутации.

Кроме того, особый тип наследственного материала содержится в митохондриях. В них ДНК несколько отличается от таковой в ядре (митохондриальная дезоксирибонуклеиновая кислота имеет кольцевидную форму и выполняет несколько другие функции).
Сама молекула может выделяться из любых клеток организма (для исследования чаще всего используют мазок с внутренней стороны щеки либо кровь). Отсутствует генетический материал только в отшелушивающемся эпителии и некоторых клетках крови (эритроцитах).
Функции
Состав молекулы ДНК обуславливает выполнение ею функции передачи информации из поколения в поколение. Это происходит за счет синтеза определенных белков, обуславливающих проявление того или иного генотипического (внутреннего) или фенотипического (внешнего - например, цвет глаз или волос) признака.

Передача информации осуществляется за счет реализации ее из генетического кода. На основании сведений, зашифрованных в генетическом коде, происходит выработка специфических информационных, рибосомальных и транспортных РНК. Каждая из них отвечает за определенное действие - информационная РНК используется для синтеза белков, рибосомальная участвует в сборке белковых молекул, а транспортная образует соответствующие белки.
Любой сбой в их работе или изменение структуры приводят к нарушению выполняемой функции и появлению нетипичных признаков (мутаций).
ДНК-тест на отцовство позволяет определить наличие родственных признаков между людьми.
Генетические тесты
Для чего в настоящее время может использоваться исследование генетического материала?
Анализ ДНК используется для определения многих факторов или изменений в организме.
В первую очередь исследование позволяет определить наличие врожденных, передающихся по наследству заболеваний. К таким болезням можно отнести синдром Дауна, аутизм, синдром Марфана.
Для определения родственных связей также можно исследовать ДНК. Тест на отцовство уже давно получил широкое распространение во многих, в первую очередь юридических, процессах. Данное исследование назначают при определении генетического родства между внебрачными детьми. Часто этот тест сдают претенденты на наследство при возникновении вопросов со стороны органов власти.
Публикации по теме
-
 Почвы среднего поволжья и их плодородие Верны ли утверждения
Почвы среднего поволжья и их плодородие Верны ли утверждения
Поволжье обладает разнообразными природными ресурсами. Из многообразия ресурсной базы района особо выделяются агроклиматические ресурсы,...
-
 Брянск, бгту: проходные баллы, группы направлений и специальностей Бгту какие предметы сдавать
Брянск, бгту: проходные баллы, группы направлений и специальностей Бгту какие предметы сдавать
УСЛОВИЯ ПРОВЕДЕНИЯ КОНКУРСА В связи с высокой востребованностью экономикой РБ специальностей: 1-36 05 01 Машины и оборудование лесного...